การค้นคว้าที่เกี่ยวข้องกับกระบวนการสังเคราะห์ด้วยแสง
12.1 การค้นคว้าที่เกี่ยวข้องกับกระบวนการสังเคราะห์ด้วยแสง
การค้นคว้าที่เกี่ยวข้องกับการสังเคราะห์ด้วยแสง
ในปี พ.ศ. 2191 ( ค.ศ. 1648 ) ได้มีการพิมพ์ผลงานของ ฌอง แบบติสท์ แวน เฮลม
องท์ (Jean Baptiste Van Helmont) นักวิทยาศาสตร์ชาวเบลเยียม ที่ทำการทดลอง
สิ่งที่น่าสนใจและมีความสำคัญมากทางชีววิทยา โดยปลูกต้นหลิวหนัก 5 ปอนด์ใน
ถังใบใหญ่ที่บรรจุดินซึ่งทำให้แห้งสนิทหนัก 200 ปอนด์แล้วปิดฝาถัง ระหว่างทำการ
ทดลองได้รดน้ำต้นหลิวที่ปลูกไว้ทุกๆ วันด้วยน้ำฝนเป็นระยะเวลา 5 ปี ต้นหลิวเจริญ
เติบโตขึ้นมาก เมื่อนำต้นหลิวที่ไม่มีดินติดอยู่ที่รากไปชั่งน้ำหนัก ปรากฏว่าต้นหลิว
หนัก 169 ปอนด์ 3 ออนซ์ และเมื่อนำดินในถัง
ไปทำให้แห้งแล้วนำไปชั่งปรากฏว่ามีน้ำหนักน้อยกว่าดินที่ใช้ก่อนทำการทดลอง
เพียง 2 ออนซ์เท่านั้น

12-1 การทดลองของฌอง แบบติสท์ แวน เฮลมองท์
ในปี พ.ศ. 2315 ( ค.ศ. 1772 ) โจเซฟ พริสต์ลีย์ (Joseph Priestley) นัก
วิทยาศาสตร์ชาวอังกฤษ ได้พิมพ์ผลงานที่ทำการทดลองโดยจ่อเทียนไขไว้
ในครอบแก้ว ปรากฎว่าสักครู่เทียนไขก็ดับ และเมื่อใส่หนูเข้าไปในครอบแก้ว
ครู่ต่อมาหนูก็ตายเมื่อนำหนูที่มีชีวิตไปไว้ในครอบแก้วเดิมที่เทียนไขดับ
ปรากฏว่า หนูตายเกือบทันที
และเมื่อจุดเทียนไขแล้วนำไปใส่ในครอบแก้วเดิมที่หนูตายอยู่แล้ว ปรากฏว่า
เทียนไขดับเกือบทันที
เขาได้ทดลองนำหนูใส่ไว้ในครอบแก้วเดียวกันกับพืชสีเขียว ปรากฎว่าทั้งพืชและหนูสามารถมี
ชีวิตอยู่ได้

12-2 การทดลองของโจเซฟ พริสต์ลีย์
พริสต์ลีย์ได้นำเอาพืชสีเขียวใส่ในครอบแก้วที่เคยจุดเทียนไขเอาไว้ก่อน
แล้ว อีก 10 วันต่อมา เมื่อ จุดเทียนไขในครอบแก้วนั้นใหม่ ปรากฏว่า เทียนไขลุกไหม้
อยู่ได้ระยะหนึ่ง โดยไม่ดับทันทีพริสต์ลีย์จึงทำการทดลองเพิ่มเติม โดยแบ่งอากาศ
หลังจากเทียนไขดับแล้วออกเป็น 2 ส่วนนำพืชใส่ไว้ในส่วนหนึ่ง และอีกส่วนหนึ่งใส่
แต่แก้วบรรจุน้ำ ทิ้งไว้ระยะหนึ่งจุดเทียนไขอากาศทั้ง 2 ส่วน พบว่าเทียนไขลุกไหม้
ได้ระยะหนึ่งในอากาศส่วนแรก แต่จะดับทันทีในอากาศส่วน
ที่สอง หลังจากนั้นเขาได้ศึกษาคุณสมบัติของก๊าซและอากาศ และทราบว่า"อากาศ
ดี" ช่วยในการเผาไหม้และการหายใจของสัตว์ แต่การหายใจของสัตว์และการเผา
ไหม้ของเทียนไขทำให้เกิด "อากาศเสีย"

12-3 การทดลองเพิ่มเติมของโจเซฟ พริสต์ลีย์
พ.ศ. 2325 (ค.ศ.1782) ฌอง ซีนีบิเยร์ (Jean Senebier) ค้นพบว่าแก๊สที่เกิด
จากการลุกไหม้ และแก๊สที่เกิดจากการหายใจของสัตว์เป็นแก๊ส
คาร์บอนไดออกไซด์ ส่วนแก๊สที่ช่วยในการลุกไหม้และแก๊สที่ใช้ในการหายใจ
ของสัตว์คือแก๊สออกซิเจนต่อมาแจน อินเก็น ฮูซ เสนอว่า พืชเก็บธาตุ
คาร์บอน ซึ่งได้มาจากแก๊สคาร์บอนไดออกไซด์ไว้ในรูปของสารอินทรีย์
ในปี ค.ศ. 1799 แจน อินเก็น-ฮูซ (Jan Ingen-Housz) นักวิทยาศาสตร์ชาวดัทช์
ได้ทำการ
ทดลองคล้ายกับโจเซฟ พริสต์ลีย์ โดยใส่พืชไว้ในครอบแก้ว แต่แยกเป็นส่วนต่างๆ
ของพืช
เช่น ลำต้น ใบ เป็นต้น แล้วทิ้งไว้ในที่มืดชั่วระยะเวลาหนึ่ง หลังจากนั้นจึงจุด
เทียนไข
ไว้ใน
ครอบแก้วแต่ละอัน พบว่าเทียนไขในครอบแก้วทุกอันไม่ติดไฟ และเมื่อ
ทำการ
ทดลองอีกครั้ง
โดยนำครอบแก้วทุกอันไปไว้ในบริเวณที่มีแสงสว่าง ระยะเวลา
หนึ่ง หลังจากนั้นจึงจุด
เทียนไข
ในครอบแก้วแต่ละอัน พบว่าในครอบแก้วที่มีส่วนของ
พืชซึ่งมีสีเขียวสามารถจุด
เทียนไขให้
ติดไฟได้ จากการทดลองดังกล่าวแจน อินเก็น-
ฮูซได้ให้ข้อสรุปไว้ว่า ส่วนของพืชที่มี
สีเขียว
สามารถเปลี่ยนอากาศเสียให้เป็นอากาศ
ดีได้ โดยพืชต้องอาศัยแสงเป็นปัจจัยใน
กระบวนการ
ดังกล่าวด้วย
 12-4 การทดลองของแจน อินเก็น-ฮูซ
12-4 การทดลองของแจน อินเก็น-ฮูซ
ในปี ค.ศ. 1804 นิโคลาส ธีโอดอร์ เดอ โซซูร์ (Nicolas Theodore de Soussure) ได้
ทำการรวบรวมและศึกษาผลงานของนักวิทยาศาสตร์ในอดีตหลายๆ ท่าน โดยอาศัย
ความรู้พื้นฐานทางด้านเคมีสมัยใหม่ ทำให้ได้ข้อสรุปเกี่ยวกับกระบวนการสังเคราะห์
ด้วยแสงของพืชดังต่อไปนี้
- พืชจะคายแก๊สคาร์บอนไดออกไซด์และแก๊สออกซิเจนในเวลากลางวัน และจะคาย
เฉพาะ
แก๊สคาร์บอนไดออกไซด์ในเวลากลางคืน แสดงว่าพืชหายใจตลอดเวลา แต่พืชมี
การสังเคราะห์แสงเฉพาะเวลากลางวันหรือเมื่อได้รับแสง
- แร่ธาตุในดินมีความจำเป็นต่อการเจริญเติบโตของพืช
- น้ำไม่ใช่เพียงละลายแร่ธาตุในดินให้แก่พืชเท่านั้น แต่น้ำยังมีบทบาทสำคัญ
โดยตรงใน
กระบวนการสังเคราะห์ด้วยแสงของพืช
พ.ศ.2405 (ค.ศ.1862) จูเลียส ซาซ (Julius Sachs) พบว่าสารอินทรีย์ที่พืช
สร้าง คือ
น้ำตาล ซึ่งเป็นสารคาร์โบไฮเดรต
ในเวลาต่อมานักวิทยาศาสตร์ได้เรียกกระบวนการสร้างคาร์โบไฮเดรตของพืช
ที่อาศัย
แสงนี้ว่า กระบวนการสังเคราะห์ด้วยแสง (photosynthesis)
พ.ศ.2438 (ค.ศ.1895) เองเกลมัน (T.W.Engelmann) ได้ทำการทดลองโดยใช้
ปริซึม
เพื่อแยกแสงออกเป็นสเปกตรัมให้แก่สาหร่ายสไปโรไจราซึ่งเจริญอยู่ในน้ำที่มี
แบคทีเรีย จากการทดลองพบว่า แบคทีเรียที่ต้องการออกซิเจนมารวมกลุ่มกัน
ที่
บริเวณสาหร่ายได้รับแสงสีแดงและสีน้ำเงินเพราะทั้งสองบริเวณนี้สาหร่ายจะ
ให้แก๊ส
ออกซิเจนมากกว่าในบริเวณอื่น
พ.ศ.2473 (ค.ศ.1973) แวน นีล (Van Niel) นักวิทยาศาสตร์ชาวสหรัฐอเมริกา
แห่ง
มหาวิทยาลัยสแตนฟอร์ดทดลองเลี้ยงแบคทีเรียที่สังเคราะห์ด้วยแสงโดยไม่
ใช้น้ำแต่
ใช้ไฮโดรเจนซัลไฟด์แทนพบว่า ผลที่ได้จากการสังเคราะห์ด้วยแสงแทนที่จะ
เกิดแก๊ส
ออกซิเจนกลับเกิดซัลเฟอร์ขึ้นแทน
พ.ศ.2484 (ค.ศ.1941) แซม รูเบน(Sam Ruben) และมาร์ติน คาเมน (Martin
Kamen)ได้ทำการทดลองใช้น้ำที่ประกอบด้วย 18O

12-5 การสังเคราะห์ด้วยแสงของพืชจะปล่อยออกซิเจนออกมา
ในปี พ.ศ. 2475 (ค.ศ. 1932) โรบิน ฮิลล์ (Robin Hill) ทำการทดลองผ่านแสง
เข้าไปในของผสมซึ่งมีเกลือเฟอริกและคลอโรพลาสต์ที่สกัดออกมาจากผัก
โขม ปรากฏว่า เกลือเฟอริกเปลี่ยนเป็นเกลือเฟอรัสและมีออกซิเจนเกิดขึ้น แต่
ถ้าในของผสมไม่มีเก
ลือเฟอริกก็จะไม่เกิดแก๊สออกซิเจน

12-6 การทดลองของ โรบิน ฮิลล์
ในปี พ.ศ. 2494 (ค.ศ. 1951) แดเนียล อาร์นอน (Daniel Arnon ) และคณะแห่ง
มหาวิทยาลัยแคลิฟอร์เนียที่เบิร์กเลย์ ได้ศึกษารายละเอียดเกี่ยวกับการ
ทดลองของ
ฮิลล์ อาร์นอน และได้ทำการทดลองดังนี้
- การทดลองที่ 1 เมื่อให้แสงแต่ไม่ให้คาร์บอนไดออกไซด์

12-7 การทดลองที่1 ของ แดเนียล อาร์นอน
- การทดลองที่ 2 เมื่อไม่ให้แสงแต่มีการเติมแก๊ส
- คาร์บอนไดออกไซด์ ATP และNADPH

การค้นคว้าที่เกี่ยวข้องกับการสังเคราะห์ด้วยแสง
ในปี พ.ศ. 2191 ( ค.ศ. 1648 ) ได้มีการพิมพ์ผลงานของ ฌอง แบบติสท์ แวน เฮลม
องท์ (Jean Baptiste Van Helmont) นักวิทยาศาสตร์ชาวเบลเยียม ที่ทำการทดลอง
สิ่งที่น่าสนใจและมีความสำคัญมากทางชีววิทยา โดยปลูกต้นหลิวหนัก 5 ปอนด์ใน
ถังใบใหญ่ที่บรรจุดินซึ่งทำให้แห้งสนิทหนัก 200 ปอนด์แล้วปิดฝาถัง ระหว่างทำการ
ทดลองได้รดน้ำต้นหลิวที่ปลูกไว้ทุกๆ วันด้วยน้ำฝนเป็นระยะเวลา 5 ปี ต้นหลิวเจริญ
เติบโตขึ้นมาก เมื่อนำต้นหลิวที่ไม่มีดินติดอยู่ที่รากไปชั่งน้ำหนัก ปรากฏว่าต้นหลิว
หนัก 169 ปอนด์ 3 ออนซ์ และเมื่อนำดินในถัง
ไปทำให้แห้งแล้วนำไปชั่งปรากฏว่ามีน้ำหนักน้อยกว่าดินที่ใช้ก่อนทำการทดลอง
เพียง 2 ออนซ์เท่านั้น
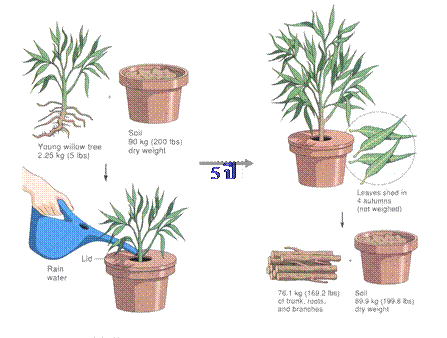
12-1 การทดลองของฌอง แบบติสท์ แวน เฮลมองท์
ในปี พ.ศ. 2315 ( ค.ศ. 1772 ) โจเซฟ พริสต์ลีย์ (Joseph Priestley) นัก
วิทยาศาสตร์ชาวอังกฤษ ได้พิมพ์ผลงานที่ทำการทดลองโดยจ่อเทียนไขไว้
ในครอบแก้ว ปรากฎว่าสักครู่เทียนไขก็ดับ และเมื่อใส่หนูเข้าไปในครอบแก้ว
ครู่ต่อมาหนูก็ตายเมื่อนำหนูที่มีชีวิตไปไว้ในครอบแก้วเดิมที่เทียนไขดับ
ปรากฏว่า หนูตายเกือบทันที
และเมื่อจุดเทียนไขแล้วนำไปใส่ในครอบแก้วเดิมที่หนูตายอยู่แล้ว ปรากฏว่า
เทียนไขดับเกือบทันที
ชีวิตอยู่ได้

12-2 การทดลองของโจเซฟ พริสต์ลีย์
พริสต์ลีย์ได้นำเอาพืชสีเขียวใส่ในครอบแก้วที่เคยจุดเทียนไขเอาไว้ก่อน
แล้ว อีก 10 วันต่อมา เมื่อ จุดเทียนไขในครอบแก้วนั้นใหม่ ปรากฏว่า เทียนไขลุกไหม้
อยู่ได้ระยะหนึ่ง โดยไม่ดับทันทีพริสต์ลีย์จึงทำการทดลองเพิ่มเติม โดยแบ่งอากาศ
หลังจากเทียนไขดับแล้วออกเป็น 2 ส่วนนำพืชใส่ไว้ในส่วนหนึ่ง และอีกส่วนหนึ่งใส่
แต่แก้วบรรจุน้ำ ทิ้งไว้ระยะหนึ่งจุดเทียนไขอากาศทั้ง 2 ส่วน พบว่าเทียนไขลุกไหม้
ได้ระยะหนึ่งในอากาศส่วนแรก แต่จะดับทันทีในอากาศส่วน
ที่สอง หลังจากนั้นเขาได้ศึกษาคุณสมบัติของก๊าซและอากาศ และทราบว่า"อากาศ
ดี" ช่วยในการเผาไหม้และการหายใจของสัตว์ แต่การหายใจของสัตว์และการเผา
ไหม้ของเทียนไขทำให้เกิด "อากาศเสีย"

12-3 การทดลองเพิ่มเติมของโจเซฟ พริสต์ลีย์
พ.ศ. 2325 (ค.ศ.1782) ฌอง ซีนีบิเยร์ (Jean Senebier) ค้นพบว่าแก๊สที่เกิด
จากการลุกไหม้ และแก๊สที่เกิดจากการหายใจของสัตว์เป็นแก๊ส
คาร์บอนไดออกไซด์ ส่วนแก๊สที่ช่วยในการลุกไหม้และแก๊สที่ใช้ในการหายใจ
ของสัตว์คือแก๊สออกซิเจนต่อมาแจน อินเก็น ฮูซ เสนอว่า พืชเก็บธาตุ
คาร์บอน ซึ่งได้มาจากแก๊สคาร์บอนไดออกไซด์ไว้ในรูปของสารอินทรีย์
ในปี ค.ศ. 1799 แจน อินเก็น-ฮูซ (Jan Ingen-Housz) นักวิทยาศาสตร์ชาวดัทช์
ได้ทำการ
ทดลองคล้ายกับโจเซฟ พริสต์ลีย์ โดยใส่พืชไว้ในครอบแก้ว แต่แยกเป็นส่วนต่างๆ
ของพืช
เช่น ลำต้น ใบ เป็นต้น แล้วทิ้งไว้ในที่มืดชั่วระยะเวลาหนึ่ง หลังจากนั้นจึงจุด
เทียนไข
ไว้ใน
ครอบแก้วแต่ละอัน พบว่าเทียนไขในครอบแก้วทุกอันไม่ติดไฟ และเมื่อ
ทำการ
ทดลองอีกครั้ง
โดยนำครอบแก้วทุกอันไปไว้ในบริเวณที่มีแสงสว่าง ระยะเวลา
หนึ่ง หลังจากนั้นจึงจุด
เทียนไข
ในครอบแก้วแต่ละอัน พบว่าในครอบแก้วที่มีส่วนของ
พืชซึ่งมีสีเขียวสามารถจุด
เทียนไขให้
ติดไฟได้ จากการทดลองดังกล่าวแจน อินเก็น-
ฮูซได้ให้ข้อสรุปไว้ว่า ส่วนของพืชที่มี
สีเขียว
สามารถเปลี่ยนอากาศเสียให้เป็นอากาศ
ดีได้ โดยพืชต้องอาศัยแสงเป็นปัจจัยใน
กระบวนการ
ดังกล่าวด้วย

12-4 การทดลองของแจน อินเก็น-ฮูซ
ในปี ค.ศ. 1804 นิโคลาส ธีโอดอร์ เดอ โซซูร์ (Nicolas Theodore de Soussure) ได้
ทำการรวบรวมและศึกษาผลงานของนักวิทยาศาสตร์ในอดีตหลายๆ ท่าน โดยอาศัย
ความรู้พื้นฐานทางด้านเคมีสมัยใหม่ ทำให้ได้ข้อสรุปเกี่ยวกับกระบวนการสังเคราะห์
ด้วยแสงของพืชดังต่อไปนี้
- พืชจะคายแก๊สคาร์บอนไดออกไซด์และแก๊สออกซิเจนในเวลากลางวัน และจะคาย
เฉพาะ
แก๊สคาร์บอนไดออกไซด์ในเวลากลางคืน แสดงว่าพืชหายใจตลอดเวลา แต่พืชมี
การสังเคราะห์แสงเฉพาะเวลากลางวันหรือเมื่อได้รับแสง
- แร่ธาตุในดินมีความจำเป็นต่อการเจริญเติบโตของพืช
- น้ำไม่ใช่เพียงละลายแร่ธาตุในดินให้แก่พืชเท่านั้น แต่น้ำยังมีบทบาทสำคัญ
โดยตรงใน
กระบวนการสังเคราะห์ด้วยแสงของพืช
พ.ศ.2405 (ค.ศ.1862) จูเลียส ซาซ (Julius Sachs) พบว่าสารอินทรีย์ที่พืช
สร้าง คือ
น้ำตาล ซึ่งเป็นสารคาร์โบไฮเดรต
ในเวลาต่อมานักวิทยาศาสตร์ได้เรียกกระบวนการสร้างคาร์โบไฮเดรตของพืช
ที่อาศัย
แสงนี้ว่า กระบวนการสังเคราะห์ด้วยแสง (photosynthesis)
พ.ศ.2438 (ค.ศ.1895) เองเกลมัน (T.W.Engelmann) ได้ทำการทดลองโดยใช้
ปริซึม
เพื่อแยกแสงออกเป็นสเปกตรัมให้แก่สาหร่ายสไปโรไจราซึ่งเจริญอยู่ในน้ำที่มี
แบคทีเรีย จากการทดลองพบว่า แบคทีเรียที่ต้องการออกซิเจนมารวมกลุ่มกัน
ที่
บริเวณสาหร่ายได้รับแสงสีแดงและสีน้ำเงินเพราะทั้งสองบริเวณนี้สาหร่ายจะ
ให้แก๊ส
ออกซิเจนมากกว่าในบริเวณอื่น
พ.ศ.2473 (ค.ศ.1973) แวน นีล (Van Niel) นักวิทยาศาสตร์ชาวสหรัฐอเมริกา
แห่ง
มหาวิทยาลัยสแตนฟอร์ดทดลองเลี้ยงแบคทีเรียที่สังเคราะห์ด้วยแสงโดยไม่
ใช้น้ำแต่
ใช้ไฮโดรเจนซัลไฟด์แทนพบว่า ผลที่ได้จากการสังเคราะห์ด้วยแสงแทนที่จะ
เกิดแก๊ส
ออกซิเจนกลับเกิดซัลเฟอร์ขึ้นแทน
พ.ศ.2484 (ค.ศ.1941) แซม รูเบน(Sam Ruben) และมาร์ติน คาเมน (Martin
Kamen)ได้ทำการทดลองใช้น้ำที่ประกอบด้วย 18O

12-5 การสังเคราะห์ด้วยแสงของพืชจะปล่อยออกซิเจนออกมา
ในปี พ.ศ. 2475 (ค.ศ. 1932) โรบิน ฮิลล์ (Robin Hill) ทำการทดลองผ่านแสง
เข้าไปในของผสมซึ่งมีเกลือเฟอริกและคลอโรพลาสต์ที่สกัดออกมาจากผัก
โขม ปรากฏว่า เกลือเฟอริกเปลี่ยนเป็นเกลือเฟอรัสและมีออกซิเจนเกิดขึ้น แต่
ถ้าในของผสมไม่มีเก
ลือเฟอริกก็จะไม่เกิดแก๊สออกซิเจน

12-6 การทดลองของ โรบิน ฮิลล์
ในปี พ.ศ. 2494 (ค.ศ. 1951) แดเนียล อาร์นอน (Daniel Arnon ) และคณะแห่ง
มหาวิทยาลัยแคลิฟอร์เนียที่เบิร์กเลย์ ได้ศึกษารายละเอียดเกี่ยวกับการ
ทดลองของ
ฮิลล์ อาร์นอน และได้ทำการทดลองดังนี้
- การทดลองที่ 1 เมื่อให้แสงแต่ไม่ให้คาร์บอนไดออกไซด์

12-7 การทดลองที่1 ของ แดเนียล อาร์นอน
- การทดลองที่ 2 เมื่อไม่ให้แสงแต่มีการเติมแก๊ส
- คาร์บอนไดออกไซด์ ATP และNADPH

{kind=link}
การสังเคราะห์ด้วยแสง(Photosynthesis)
กระบวนการสังเคราะห์ดว้ยแสง เป็นกระบวนการที่สิ่งมีชีวิตเปลี่ยนพลงังานแสงเป็นพลงังานเคมีที่เกบ็
สะสมไวใ้นสารอินทรียท์ ี่สิ่งมีชีวิตสร้างข้ึน และพลงังานเคมีที่สร้างข้ึนน้ีจดัว่าเป็นแหล่งพลงังานที่สิ่งมีชีวิตต่างๆ
ในโลกน้ีไดใ้ชป้ระโยชน์
คุณสมบัติของแสง
แสงมีคุณสมบัติเป็ นคลื่นแม่เหล็กไฟฟ้ า ช่วงความยาวคลื่นแสงที่เรามองเห็นได้ หรือ visible light มี
ช่วงความยาวคลื่นประมาณ 400 ถึง 700 nm (ดังภาพข้างล่าง) ค่าความยาวคลื่น ( หรือระยะการเคลื่อนที่
ของ 1 ช่วงคลื่นเป็ นสมบัติที่ส าคัญของคลื่น และสัมพันธ์กับความถี่ของคลื่น ซึ่งหมายถึงจ านวนช่วงคลื่นที่
เคลื่อนที่ผ่านจุดจุดหนึ่งต่อหน่วยเวลา ความยาวคลื่นแปรผันกลับกับความถี่ โดย
c = โดย c = ความเร็วของคลื่น
แสงยังมีคุณสมบัติที่ส าคัญอีกประการหนึ่งคือ เป็ นอนุภาค หรือ photon แต่ละ photon มีหน่วย
พลังงานเรียกว่า quantum พลงังานแสงแต่ละอนุภาคข้ึนกบัความถี่ของแสง ตามกฎของ Planck ดังสมการ
E= h โดย h = Plank's constant = 6.626x 10-34 Js
ตามสมการท้งัสองขา้งตน้ สามารถประมวลไดว้่าหากค่าความยาวคลื่นเพิ่มสูงข้ึน พลงังานต่อ photon
ก็จะลดลง ตัวอย่างเช่น อนุภาคของแสงสีม่วงมีพลังงานมากกว่าอนุภาคของแสงสีแดง
สิ่งมีชีวิตที่สามารถสังเคราะห์ด้วยแสงได้
สิ่งมีชีวิตที่สามารถสังเคราะห์ดว้ยแสงได้มีท้งั prokaryote ได้แก่ photosynthetic bacteria
และ cyanobacteria และ eukaryote ไดแ้ก่สิ่งมีชีวิตในกลุ่มสาหร่าย มอส เฟิร์น สน ปรงและพืชดอกเหล่า
สิ่งมีชีวิตที่สามารถสังเคราะห์ดว้ยแสงไดน้้ีมีpigment หรือรงควัตถุที่แตกต่างกันไป (ดังแสดงในตารางที่ 1)
เพื่อใช้ในการรับพลังงานแสงเพื่อน ามาใช้ในกระบวนการสังเคราะห์ด้วยแสง
ตารางที่ 1 แสดงชนิดของรงควัตถุที่ใช้ในการสังเคราะห์ด้วยแสงในสิ่งมีชีวิตต่าง ๆ

รงควัตถุที่ใช้รับพลังงานแสง
การที่สิ่งมีชีวิตสามารถสร้างพนัธะเคมีในโมเลกุลของสารอินทรียไ์ดโ้ดยกระบวนการสังเคราะห์ดว้ย
แสงน้นั สิ่งมีชีวิตตอ้งมีโมเลกุลที่มีความสามารถในการดูดกลืนแสงโมเลกุลดงักล่าวคือ pigment หรือรงค
วัตถุ ซึ่งมีคุณสมบัติเป็ น photoreceptor นนั่ เอง
pigment ที่สิ่งมีชีวิตใชใ้นกระบวนการสังเคราะห์ดว้ยแสงสามารถแบ่งตามลกัษณะโครงสร้างของ
โมเลกุลเป็ น 3 ประเภท คือ
1. Chlorophyll และ Bacteriochlorophyll
Chlorophyllและ Bacteriochlorophyll เป็ น pigment ที่พบไดท้วั่ ไปในสิ่งมีชีวิตที่สามารถ
สังเคราะห์ด้วยแสงได้ โดย pigment ในกลุ่มน้ีมีมีโครงสร้างที่ประกอบดว้ย porphyrin-like
structure มี Mg2+ อยู่ตรงกลางของโครงสร้าง และมีส่วนที่เป็ นสายยาวของไฮโดรคาร์บอน ซึ่งเป็ น
hydrophobic region ซึ่งฝังอยู่บริเวณ photosynthetic membrane

2. Phycobilins หรือ Bilin pigments
Phycobilins หรือ Bilin pigments เป็ น pigment ที่พบมากใน cyanobacteria และสาหร่าย
สีแดง มีโครงสร้างที่เรียกว่า open-chain tetrapyrroles Phycobilins ที่เกี่ยวข้องกับกระบวนการ
สังเคราะห์ด้วยแสงสามารถแบ่งได้เป็ น 3 ชนิด คือ phycoerythrin (phycoerythrobilin)
phycocyanin (phycocyanobilin) และ allophycocyanin (allophycocyanobilin)

3. Carotenoids
Carotenoids เป็ น pigment ที่พบทวั่ ไปในสิ่งมีชีวิตที่สามารถสังเคราะห์ดว้ยแสงได้มีหนา้ที่ในการ
ช่วยรับพลังงานแสง เพื่อน ามาใช้ในกระบวนการสังเคราะห์ด้วยแสง และยังมีความสามารถในการป้ องกัน
อันตรายจากแสง
Carotenoids มีโครงสร้างหลักเป็ นสายไฮโดรคาร์บอน ที่ประกอบด้วยคาร์บอน 40 อะตอม สามารถ
จ าแนกได้เป็ น 2 กลุ่มคือ carotenes และ xanthophylls

การดูดกลืนแสง
pigment แต่ละชนิดมีคุณสมบตัิเฉพาะตวัที่แตกต่างกนัดงัที่กล่าวไปแลว้ คุณสมบตัิที่แตกต่างกนั น้ีทา
ให้ pigment แต่ละชนิดมีความสามารถในการดูดกลืนแสงในช่วงคลื่นที่ต่างกันด้วยดังแสดงในภาพข้างล่าง
น้ี
ในสิ่งมีชีวิตที่เป็น prokaryote pigment ที่ใช้ในกระบวนการสังเคราะห์ด้วยแสงจะฝังตัน

ในสิ่งมีชีวิตที่เป็น prokaryote pigment ที่ใช้ในกระบวนการสังเคราะห์ด้วยแสงจะฝังตัวอยู่บน
photosynthetic membrane ในขณะที่ใน eukaryote จะมีคลอโรพลาสต์ ซึ่งเป็ นออร์กาเนลล์เฉพาะที่ท า
หน้าที่ในการสังเคราะห์ด้วยแสง
คลอโรพลาสต์
กระบวนการสังเคราะห์ดว้ยแสงในพืชเกิดข้ึนที่คลอโรพลาสต์คลอโรพลาสตเ์ป็นออร์กาเนลลท์ ี่มีเยอื่
หุม้ สองช้นัคือเยอื่ หุม้ ช้นั นอก (outer membrane) และเยอื่ หุม้ ช้นั ใน(inner membrane) มี DNA
RNA และ ribosome ของตัวเอง สามารถสังเคราะห์โปรตีนได้เอง ของเหลวที่บรรจุอยู่ในคลอโรพลาสต์
ส่วนใหญ่เป็นเอนไซมท์ ี่ใชใ้นการสังเคราะห์ดว้ยแสง เรียกส่วนที่มีของเหลวบรรจุอยนู่ ้ีว่าstroma ส่วน
thylakoid membrane มีลกัษณะเป็นเมมเบรนที่พบั ซ้อนทบักนัไปมาเป็นถุงแบนซ้อนเป็นต้งัๆ เรียกว่า
grana หรือ grana lamellae โดยแต่ละต้งัเรียกว่า granum ภายในถุงแต่ละถุงที่เรียงเป็นต้งัมีของเหลวอยู่เอกสารประกอบการสอนรายวิชา 2305101 GEN BIO II ภาคปลาย ปี การศึกษา 2551
ผศ.ดร.กนกวรรณ เสรีภาพ
เรียกส่วนที่อยภู่ ายในถุงน้ีว่า lumen หรือ thylakoid space บริเวณ thylakoid membrane น้ีเองที่เป็นที่
อยู่ของ pigments ที่ใช้ในการตรึงพลังงานแสง
pigments ต่างๆ ที่อยู่ที่ บริเวณ thylakoid membrane มีการจัดเรียงตัวอย่างเป็ นระบบเพื่อให้การ
ตรึงพลังงานแสงมีประสิทธิภาพสูงที่สุด โดยการจัดเรียงตัวของ pigmentsอยา่ งเป็นระบบน้ีเรียกว่า antenna
complex ประกอบด้วย แคโรทีนอยด์ คลอโรฟิ ลล์ บี และคลอโรฟิ ลล์ เอ ท างานร่วมกันในการรับพลังงานแสง
แลว้ส่งพลงังานน้นั เขา้สู่ศูนยก์ลางปฏิกิริยา (reaction center) ซึ่งคือโมเลกุลของคลอโรฟิ ลล์ เอ2โมเลกุล
โมเลกุลคลอโรฟิ ลล์ เอ2 โมเลกุลน้ีเมื่อไดร้ับพลงังานในช่วงคลื่นที่เหมาะสม อิเล็คตรอนในโมเลกุลของ
คลอโรฟิ ลล์ เอ 2 โมเลกุลน้ีจะถูกกระตุน้ ใหม้ีพลงังานสูงข้ึนจากภาวะเดิมที่อยู่ที่ ground state จะถูกกระตุ้นไป
อยใู่ นภาวะที่มีพลงังานสูงข้ึน คือ ภาวะ excited state พร้อมที่จะปลดปล่อยอิเลค็ ตรอนน้ีใหก้ บั
ตัวรับอิเล็คตรอนตัวถัดไป
กลไกกระบวนการสังเคราะห์ด้วยแสง
ในการเปลี่ยนพลงังานใหเ้ป็นพลงังานเคมีโดยกระบวนการสังเคราะห์ดว้ยแสงน้นั เราอาจจา แนกเป็น 2
ข้นั ตอนคือ
1. ปฏิกิริยาที่ต้องใช้แสง (Light reaction) เป็นข้นั ตอนที่เกิดข้ึนบริเวณ thylakoid รงควัตถุ
ต่างๆ ที่อยู่บริเวณ thylakoid ซึ่งอยู่รวมกันเป็ นกลุ่มโดยมีการเกาะตัวอยู่กับโปรตีนหลายชนิด
กลุ่มของโปรตีนบน thylakoid ที่มีรงควตัถุประกอบอยดู่ ว้ยน้ีเรียกว่า ระบบแสง
(photosystem) ท าหน้าที่รับพลังงานแสงมาใช้ในการสร้างสารที่มีพลังงานสูง คือ ATP และ
NADPH เพื่อน าพลังงานที่ได้ไปใช้ในการสร้างสารอินทรีย์ในกระบวนการตรึง
คาร์บอนไดออกไซด์ต่อไป
2. ปฏิกิริยาการตรึงคาร์บอนไดออกไซด์ (CO2 fixation reaction) เป็นข้นั ตอนที่เกิดข้ึนบริเวณ
stroma ซึ่งมีเอนไซม์ที่เกี่ยวข้องกับการตรึงคาร์บอนไดออกไซด์อยู่ โดยใช้พลังงานจาก ATP
และ NADPH ที่ไดจ้ากปฎิกิริยาที่ตอ้งใชแ้สงเพื่อสร้างเป็นสารอินทรียใ์นรูปของน้า ตาล
ปฏิกิริยาที่ต้องใช้แสง (Light reaction)
ในข้นั ตอนการเปลี่ยนพลงังานแสงใหเ้ป็นพลงังานเคมีในรูปของ ATP และ NADPH ต้องผ่านระบบ
การรับพลังงานแสงและกระบวนการถ่ายทอดพลังงานในรูปของการถ่ายทอดอิเลคตรอน พืชมีระบบแสง 2 ระบบ
คือ ระบบแสงI (photosystem I)และระบบแสง II (photosystem II) ซ่ึงระบบแสงท้งัสองจะทา หนา้ที่
ร่วมกันเพื่อให้สามารถถ่ายทอดพลังงานแสงไปใช้ในการสร้าง ATP และ NADPH การถ่ายทอดอิเลคตรอนใน
light reaction ระบบแสง หรือ photosystem เป็ น protein complex ที่ฝังตัวอยู่ใน thylakoid
membrane แต่ละ photosystem ประกอบด้วย หน่วยรับพลังงานแสง ซึ่งมีรงควัตถุหลายชนิดเป็ น
องค์ประกอบโปรตีนชนิดต่างๆมีความจ าเพาะต่อการท างานของระบบแสงแต่ละชนิด
ระบบแสง I ประกอบด้วยหน่วยรับพลังงานแสงที่มีศูนย์กลางปฏิกิริยาที่สามารถรับพลังงานในช่วง
คลื่นที่มีความยาวคลื่นไม่ต ่ากว่า 700 นาโนเมตร (P700) ในขณะที่ ระบบแสง II จะประกอบด้วยหน่วยรับ
พลังงานแสงที่มีศูนย์กลางปฏิกิริยาที่สามารถรับพลังงานในช่วงคลื่นที่มีความยาวคลื่นไม่ต ่ากว่า 680 นาโนเมตร เอกสารประกอบการสอนรายวิชา 2305101 GEN BIO II ภาคปลาย ปี การศึกษา 2551
ผศ.ดร.กนกวรรณ เสรีภาพ
(P680) ปฏิกิริยาที่ต้องใช้แสงมี pathway ที่ใช้ในการถ่ายทอดอิเลคตรอน 2 pathway คือ การถ่ายทอด
อิเลคตรอนแบบไม่เป็ นวัฏจักร (non-cyclic electron transfer) และ การถ่ายทอดอิเลคตรอนแบบเป็ นวัฏ
จักร (cyclic electron transfer)
การถ่ายทอดอิเลคตรอนแบบไม่เป็ นวัฏจักร (non-cyclic electron transfer)

จากภาพ เมื่อ antenna complex ได้รับแสงและส่งทอดอิเลคตรอนต่อให้ก ับ reaction centerคือ
ระบบแสง II แล้ว ระบบแสง II จะส่งอิเลคตรอนต่อไปให้กับตัวรับอิเลคตรอน ท าให้ระบบแสง II มี
ความสามารถในการช่วงชิงอิเลคตรอนเพิ่มข้ึน (เป็ น strong oxidant) ระบบแสง II จะสามารถ
ดึงอิเลคตรอนมาจากโมเลกุลของน้า ได้ปฏิกิริยาการแยกโมเลกุลของน้า ดว้ยพลงังานแสงน้ีเรียกว่า photolysis
ในปฏิกิริยา photolysis น้ีจะเกิด O2 และ H
+
เกิดข้ึน โดย H
+
ที่เกิดข้ึนน้ีจะสะสมอยใู่ น thylakoid space
หรือ lumen
ส่วนตัวรับอิเลคตรอนเมื่อได้รับอิเลคตรอนจากระบบแสง IIแล้ว ก็จะส่งทอดอิเลคตรอนต่อไปให้กับ
ตัวรับอิเลคตรอนอื่นๆ ใน ระบบถ่ายทอดอิเลคตรอน (electron transport system หรือ ETS)เช่น plasto
quinone plastocyanin cytochrome b6f complex ก็เป็ นหนึ่งในระบบ ETS โดย cytochrome
complex น้ีฝังตวัอยใู่ น thylakoid membrane เช่นกัน ระบบ ETS ช่วยให้เกิดการถ่ายเทอิเลคตรอนต่อ
ไปเป็ นทอดๆ ยังตัวรับอิเลคตรอนต่างๆ ในระบบ ในที่สุดก็จะส่งทอดอิเลคตรอนถึง ระบบแสง I ซึ่งบริเวณระบบ
แสง I น้ีกจ็ะไดร้ับพลงังานแสงมากระตุน้อิเลคตรอนใหม้ีพลงังานที่สูงข้ึนอีกคร้ังหน่ึงและสามารถถ่ายทอด
อิเลคตรอนให้กับตัวรับอิเลคตรอนตัวต่อไปได้ อย่างไรก็ตาม ระบบแสง I เมื่อสูญเสียอิเลคตรอนให้กับ
ตวัรับอิเลคตรอนแลว้ ไม่สามารถดึงอิเลคตรอนจากน้า ไดเ้หมือนที่เกิดข้ึนในระบบแสง II แต่สามารถ
รับอิเลคตรอนจากโมเลกุลอื่นๆ ที่ได้รับถ่ายทอดมาจากระบบแสง II และมีการส่งทอดอิเลคตรอนให้กับ
ตวัรับอิเลคตรอนอื่นๆ ในระบบ จนกระทงั่ ถึงตวัรับอิเลคตรอนตวัสุดทา้ยคือ NADP+
ซึ่งได้รับ 2 อิเลคตรอน
และ 1 โปรตอนและ ได้เป็ น NADPH โดยการท างานของเอนไซม์ NADP+
reductase
ส่วนในกระบวนการสร้าง ATP หรือ photophosphorylation พืชใช้พลังงานที่ได้จากแสงมาใช้
ในกระบวนการ phosphorylation หรือการเติมหมู่ฟอสเฟตให้กับ ADP นนั่ เองโดยในระหว่างที่มีการ
ถ่ายทอดอิเลคตรอนจาก ระบบแสง II ไปยัง ระบบแสง I น้นั พลงังานบางส่วนจะถูกนา ไปใชใ้นการลา เลียง
โปรตอนจาก stroma ไปยัง thylakoid lumen รวมท้งัในข้นั ตอนการแตกตวัของน้า ยงัเกิดการสะสมของง
โปรตอน ใน thylakoid space หรือ lumen ดว้ยดงัน้นั จึงเกิดความต่างศกัยร์ะหว่างthylakoid lumen
และ stroma เนื่องจากมีความเข้มข้นของโปรตอนใน lumen สูงกว่าใน stroma ศกัยไ์ฟฟ้าที่แตกต่างกนั น้ีถูก
น าไปใช้ในการสร้าง ATP โดยการเร่งปฏิกิริยาของเอนไซม์ ATP synthase ซึ่งฝังตัวอยู่ที่ thylakoid
membrane
ดงัน้นั ในการถ่ายทอดอิเลคตรอนจากระบบแสง II ไปยังตัวรับอิเลคตรอนตัวสุดท้ายจนได้เป็ น
NADPH น้ีจึงจัดเป็ นการถ่ายทอดอิเลคตรอนแบบไม่เป็ นวัฏจักร (non-cyclic electron transfer) โดย
ในกระบวนการน้ีจะเกิดการแตกตวัของน้า ไดอ้อกซิเจน โปรตอนและอิเลคตรอน และสร้างสารพลงังานสูงคือ
ATP และ NADPH
การถ่ายทอดอิเลคตรอนแบบเป็ นวัฏจักร (cyclic electron transfer)
ส าหรับการถ่ายทอดอิเลคตรอนแบบเป็ นวัฏจักร เมื่อระบบแสง I ถ่ายทอดอิเลคตรอนไปยัง
ตัวรับอิเลคตรอนตัวถัดไปแล้ว อาจไม่ได้ส่งทอดอิเลคตรอนไปถึงตัวรับอิเลคตรอนตัวสุดท้ายก็ได้ แต่ส่งอิเลค
ตรอนกลับมายังระบบแสง II แทน ซ่ึงจากกน้นักส็ ามารถส่งอิเลคตรอนกลบัไปยงัระบบแสง Iอีกคร้ังหน่ึง เกิด
เป็นการถ่ายทอดอิเลคตรอนแบบวฏัจกัรเกิดข้ึน (cyclic electron transfer) พลังงานแสงที่ถูกถ่ายทอดใน
กรณีน้ีจะไม่ถูกนา ไปสังเคราะห์NADPH แต่สามารถน าไปใช้สร้าง ATP ได้

การถ่ายทอดอิเลคตรอนแบบเป็นวฏัจกัรน้ีอาจเกิดข้ึนไดใ้นภาวะที่พืชตอ้งการ ATP เพิ่มข้ึน หรือใน
ภาวะที่ ไม่มี NADP+
เพียงพอที่จะน าไปสร้างเป็ น NADPH
ปฏิกิริยาการตรึงคาร์บอนไดออกไซด์ (CO2 fixation reaction)
ปฏิกิริยาการตรึงคาร์บอนไดออกไซด์ หรือ calvin cycle เป็ นปฏิกิริยาที่น าพลังงานจาก ATP และ
NADPH มาใช้ในการสร้างโมเลกุลของสารอินทรีย์จากสารอนินทรีย์ แต่เดิมเชื่อว่า พืชไม่จ าเป็ นต้องใช้แสงใน
ปฏิกิริยาการตรึงคาร์บอนไดออกไซด์จึงเรียกกระบวนการน้ีว่า dark reaction แต่ต่อมาพบว่า เอนไซม์ที่ใช้ใน
กระบวนการน้ีตอ้งถูกกระตุน้ดว้ยแสงก่อน จึงสามารถทา งานได้ปัจจุบนัจึงไม่นิยมเรียกปฎิกิริยาน้ีว่า dark
reaction
ปฏิกิริยาการตรึงคาร์บอนไดออกไซด์เกิดข้ึนที่บริเวณ stroma ประกอบด้วย 3 ข้นั ตอนที่สา คญั คือ
1. Carboxylation หรือ CO2 fixation ในข้นั ตอนน้ีribulose-1,5-bisphosphate
(RuBP)รวมกับ CO2 และสร้างเป็ น 3-phosphoglycerate (3PGA) 2 โมเลกุล ซึ่งสารที่มีความ
เสถียร โดยการท างานของเอนไซม์ ribulose-1,5-bisphosphate carboxylase/oxygenase
(Rubisco)
2. Reduction ในข้นั ตอนน้ี3-phosphoglycerate ถูก reduced กลายเป็ น glyceraldehyde-
3-phosphate ซ่ึงเป็นสารประเภทน้า ตาล ATP และ NADPH จะถูกใชไ้ปในข้นั ตอนน้ี
3. Regeneration ในข้นั ตอนน้ีมีการสร้าง RuBP เพิ่อที่จะกลบัไปเป็นตวัรับ CO2 ในรอบต่อไป ใน
ข้นั ตอนน้ีอาศยัพลงังานจาก ATP ที่ได้จาก light reaction

พืช C3 พืช C4 และพืช CAM
พืช C3 เป็ นพืชที่มีการตรึงคาร์บอนไดออกไซด์ด้วย calvin cycle เพียงอย่างเดียว โดยสารตัวแรกที่
เกิดจากการตรึงคาร์บอนไดออกไซด์ คือ PGA ซึ่งเป็ นสารที่มีคาร์บอน 3 อะตอม เราจึงเรียกพืชกลุ่มน้ีว่า พืช C3
โครงสร้างภายในของพืช C3 ประกอบด้วย mesophyll cells 2 แบบคือ palisade mesophyll
และ spongy mesophyll และมีกลุ่มเน้ือเยอื่ ลา เลียงแทรกอยู่อาจมีกลุ่มเซลลท์ ี่เรียกว่า bundle sheath cell
ล้อมรอบกลุ่ม ท่อล าเลียงหรือไม่ก็ได้ และหากมี bundle sheath cell ของพืช C3 มักไม่พบคลอโรพลาสต์
การตรึงคาร์บอนไดออกไซด์ด้วย calvin cycle ของพืช C3 น้ีเกิดที่mesophyll cells เป็ นหลัก

ส่วนพืช C4 เป็ นพืชที่มี bundle sheath cell ที่มีคลอโรพลาสต์ ล้อมรอบกลุ่มท่อล าเลียง พืช C4
เป็ นพืชที่มีการตรึงคาร์บอนไดออกไซด์ 2 คร้ังโดยคร้ังแรกตรึงที่mesophyll cell โดยมีตัวมารับ
คาร์บอนไดออกไซดท์ ี่ละลายอยใู่ นน้า ในรูปของ HCO3
-
คือ phosphoenol pyruvate (PEP) ได้เป็ น
สารประกอบคาร์บอน 4 อะตอม คือ oxaloacetate โดยการท างานของเอนไซม์ PEP carboxylase ซึ่งอยู่
บริเวณ cytosol ของ mesophyll cell oxaloacetate ถูกเปลี่ยนเป็ น malate และเคลื่อนที่เข้าสู่ bundle
sheath cell ภายใน bundle sheath cell สาร C4 คือ malate จะถูกเปลี่ยนเป็ นสาร C3 และ
คาร์บอนไดออกไซด์ ในคลอโรพลาสต์ที่ bundle sheath cell คาร์บอนไดออกไซด์ จะเข้าสู่ calvin cycle
ต่อไปส่วนสาร C3 ก็จะถูกน ากลับมายัง mesophyll cell เพื่อเปลี่ยนเป็ น PEP ส าหรับการตรึง
คาร์บอนไดออกไซด์ในคร้ังต่อไป จากกลไกการตรึงคาร์บอนไดออกไซดด์งักล่าวจึงทา ใหค้วามเขม้ขน้ของ
คาร์บอนไดออกไซด์บริเวณ bundle sheath cell ของพืช C4 จึงสูงกว่าบริเวณ mesophyll cell ของพืช
C3

พืช CAM เป็ นพืชที่มีการตรึงคาร์บอนไดออกไซด์ 2 คร้ังคลา้ยคลึงกบั พืช C4 โดยมีสารอินทรียที่เป็ น
ตัวรับ HCO3
-
คือ PEP โดยการเร่งปฎิกิริยาโดยเอนไซม์ PEP carboxylase ที่ cytosol ในเวลากลางคืน
จากน้นั malate ซึ่งเปลี่ยนมาจาก oxaloacetate จะถูกน าไปเก็บไว้ที่ vacuole ในตอนกลางวันซึ่งเป็ นเวลาที่
ปากใบปิ ด malate ที่อยู่ที่ vacuoleจะถูกน ามาสลายให้ได้คาร์บอนไดออกไซด์ เพื่อน าไปใช้ในกระบวนการ
ตรึง คาร์บอนไดออกไซด์ด้วย calvin cycle อีกคร้ังที่คลอโรพลาสต์การที่ปากใบปิดในตอนกลางวนัและมี
การปลดปล่อยคาร์บอนไดออกไซด์ จาก malate ท าให้พืช CAM มีปริมาณความเข้มข้นของ
คาร์บอนไดออกไซด์สูง

Photorespiration
photorespiration เป็ นกระบวนการที่ตรึง O2 และปลดปล่อย CO2 เช่นเดียวกับกระบวนการ
หายใจ โดย O2 รวมกับ RuBP ได้ 3-phosphoglycerate และ 2-phosphoglycolate ซึ่งเป็ นเป็ นสารที่
มีคาร์บอน 3 และ 2 อะตอมตามล าดับ 3-phosphoglycerate สามารถถูกนา ไปสร้างน้า ตาลได้ส่วน 2-
phosphoglycolate จะถูกเปลี่ยนกลับเป็ น 3-phosphoglycerate ด้วยกระบวนการต่างๆ ทางชีวเคมีใน
peroxisome และ mitochondria อย่างไรก็ตามกระบวนการ photorespiration ท าให้พืชสูญเสีย
คาร์บอนในรูปของ CO2
การตรึง O2 ในกระบวนการ photorespiration อาศัยการท างานของเอนไซม์ Rubisco Rubisco
เป็นเอนไซมท์ ี่เร่งปฏิกิริยาไดท้้งัปฏิกิริยา carboxylation และ oxygenation ซ่ึงท้งัสองปฏิกิริยามีสารต้งัตน้
ตัวเดียวกันคือ RuBP โดยในการเกิด carboxylation และ oxygenation น้นั O2 ใช้ active site ของ
Rubiscoที่ต าแหน่งเดียวกับ CO2 แต่โดยปกติอัตราการเกิดปฏิกิริยาการตรึงคาร์บอนไดออกไซด์สูงกว่าการ
ตรึงออกซิเจนมากจึงทา ใหก้ระบวนการสังเคราะห์ดว้ยแสงดา เนินไดท้้งัที่ความเขม้ขน้ของคาร์บอนไดออกไซด์
ต ่ากว่าออกซิเจน
เนื่องจาก พืช C4 และพืช CAM มีกลไกในการท าให้มีความเข้มข้นของ CO2 ในเซลล์สูงกว่าพืช C3
จึงท าให้พืช C4 และพืช CAM เกิดกระบวนการ Photorespiration น้อยกว่าในพืช C3

ัจจัยบางประการที่มีผลต่ออัตราการสังเคราะห์ด้วยแสง
อตัราการสังเคราะห์ดว้ยแสงของพืช มีความสา คญั อยา่ งยงิ่ ต่อผลผลิตโดยรวมของพืช มีปัจจยัหลาย
ประการที่มีอิทธิพลต่ออตัราการสังคราะห์ดว้ยแสงของพืช ท้งัปัจจยัภายนอกและปัจจยัภายในพืช อยา่ งไรกต็าม
ในที่น้ีจะขอกล่าวถึงเฉพาะปัจจยัแวดลอ้ มภายนอกที่มีผลจา กดัหรือส่งเสริมอตัราการสังเคราะห์ดว้ยแสงของพืช
ดงัน้ี
1. แสงและความเข้มแสง แสงอาทิตย์ที่ส่องลงมายังโลก มีความเข้มแสงที่แตกต่างกันไปตามต าแหน่ง
ต่างๆ บนพ้ืนโลกและยงัแตกต่างกนั ตามฤดูกาลอีกดว้ย โดยทวั่ ไปแลว้ ความเขม้แสงที่เพิ่มสูงข้ึนมีผลทา ใหอ้ ตัรา
การสังเคราะห์ดว้ยแสงของพืชเพิ่มสูงข้ึนดว้ยแต่อยา่ งไรกต็าม อตัราการสังเคราะห์ดว้ยแสงจะเพิ่มสูงข้ึนตาม
ระดบัความเขม้ของแสงถึงจุดหน่ึงเท่าน้นั แลว้จะคงที่ณ ความเขม้แสงหน่ึง เราเรียกจุดความเขม้แสงดงักล่าวว่า
light saturation point

สา หรับ ผลของความเขม้แสงที่เพิ่มข้ึนส่งผลใหเ้กิดความแตกต่างกนั ในพืช C3
, C4 และ พืช CAM อีก
ดว้ย ดงัน้นั หากเปรียบเทียบประสิทธิภาพในการสังเคราะห์ดว้ยแสงของใบ (leaf photosynthetic capacity) ของ
พืช C3
, C4 และ พืช CAM ที่วัดในสภาวะที่มีความเข้มข้นของ CO2
และ O2 ที่ระดับปกติ อุณหภูมิพอเหมาะ
ความช้ืนสัมพทัธ์สูงและความเขม้แสงสูงจนถึงจุดlight saturation point จะพบว่า พืชที่มีอัตราการสังเคราะห์ด้วย
แสงสูงสุดส่วนใหญ่เป็ นพืช C4 และรองลงมาคือพืช C3
ส่วนพืช CAM มีอัตราการสังเคราะห์ด้วยแสงน้อยที่สุด

2. อุณหภูมิใบพืชที่ไดร้ับแสงโดยตรงจะมีผลทา ใหอุ้ณหภูมิของใบเพิ่มสูงข้ึนอยา่ งหลีกเลี่ยงไม่ได้แต่
พืชมีกลไกในการระบายความร้อนหลายวิธี เช่น การปลดปล่อยความร้อนจากใบพืชโดยตรง หรือปลดปล่อยความ
ร้อนโดยแฝงไปกบักระบวนการคายน้า ของพืช อยา่ งไรกต็ามหากพิจารณาอิทธิพลของอุณหภูมิเพียงอยา่ งเดียวต่อ
อตัราการสังเคราะห์ดว้ยแสงของพืช พบว่าอตัราการสังเคราะห์ดว้ยแสงของพืชจะเพิ่มข้ึน เมื่ออุณหภูมิสูงข้ึน
ในช่วง 0-35 °C หรือ 0-40 °C อยา่ งไรกต็าม หากอุณหภูมิสูงกว่าน้ีอตัราการสังเคราะห์ดว้ยแสงของพืชส่วนใหญ่
จะลดลง ท้งัน้ีเนื่องจากอุณหภูมิมีผลต่อกระบวนการทางชีวเคมีต่างๆ ในกระบวนการสังเคราะห์ดว้ยแสง เช่น การ
ท างานของเอนไซม์ใน Calvin cycle และ อุณหภูมิที่สูงเกินไปหรือต ่าเกินไปยังส่งผลต่อคุณสมบัติความเป็ น
semipermeability ของ cell membrane ที่จ าเป็ นต่อการท างานของกระบวนการสังเคราะห์ด้วยแสง
นอกจากน้ีผลของอุณหภูมิที่เพิ่มข้ึนจะมีความแตกต่างกนั ในพืช พืช C3
และ พืช C4
อีกด้วย โดยเมื่อ
อุณหภูมิสูงข้ึน อตัราการสังเคราะห์ดว้ยแสงของพืช C3
จะลดลง เนื่องจากการเพิ่มข้ึนของphotorespiration แต่ใน
พืช C4
ที่มี photorespiration ที่ต่า อุณหภูมิที่สูงข้ึนจึงมีผลต่ออตัราการสังเคราะห์ดว้ยแสงของพืช C4
น้อยมาก
3. ความเข้มข้นของคาร์บอนไดออกไซด์โดยทวั่ ไปแลว้ ถา้ความเขม้ขน้ของ CO2
เพิ่มข้ึน อตัราการ
สังเคราะห์ดว้ยแสงของพืชจะเพิ่มสูงข้ึนตามไปดว้ย เมื่อความเขม้ขน้ของ CO2
ในอากาศเพิ่มข้ึนจนถึงระดบั หน่ึงที่
ท าให้อัตราการตรึง CO2
ของพืชเท่ากับอัตราการปล่อย CO2 โดยกระบวนการหายใจ เราเรียกความเข้มข้นของ
CO2 ที่ระดบั น้ีว่า CO2
compensation point ซึ่งเป็ นระดับที่มีการแลกเปลี่ยนของ net CO2
เป็ นศูนย์
พืช C4
มี CO2
compensation point ที่ระดับความเข้มข้นต ่ากว่าพืช C3
มาก เนื่องจากพืช C4
มีกลไกใน
การเพิ่มความเขม้ขน้ของ CO2
ใน bundle sheet cell ดงัน้นั พืช C4
จึงต้องการ CO2
ในระดับความเข้มข้นที่ไม่มาก
นักก่อนที่จะถึง CO2
compensation point แต่ในพืช C3
ที่มี photorespiration สูง เมื่อ CO2
ในอากาศต ่า มีCO2
compensation point ในระดับที่สูงกว่า

เมื่อความเข้มข้นของ CO2 เพิ่มข้ึน อตัราการสังเคราะห์ดว้ยแสงของพืชจะเพิ่มสูงข้ึนจนถึงระดบั ที่มีCO2
อิ่มตวัในอากาศ ณ จุดดงักล่าวอตัราการตรึง CO2
จะคงที่ ส าหรับพืช C4
ซึ่งมี CO2
saturation point ต ่ากว่าพืช C3
คาร์บอนไดออกไซดจ์ะมีผลต่ออตัราการสังเคราะห์ดว้ยแสงมากนอ้ยแค่ไหนข้ึนอยกู่ บั ปัจจยัอื่นดว้ย เช่น
ความเขม้ขน้ สูงข้ึน แต่ความเขม้ของแสงนอ้ยและอุณหภูมิของอากาศกต็่า กรณีเช่นน้ีอตัราการสังเคราะห์ดว้ย
แสงจะลดต่า ลงตามไปดว้ย ในทางตรงกนัขา้ม ถา้คาร์บอนไดออกไซดม์ ีความเขม้ขน้ สูงข้ึน ความเขม้ของแสงและ
อุณหภูมิของอากาศกเ็พิ่มข้ึน กรณีเช่นน้ีอตัราการสังเคราะห์ดว้ยแสงกจ็ะสูงข้ึนตามไปดว้ย
ปัจจัยจ ากัดของการสังเคราะห์ด้วยแสง
จากปัจจยัต่างๆ ที่เกี่ยวขอ้งกบัการสังเคราะห์ดว้ยแสงตามที่กล่าวมาแลว้น้นั อาจสรุปไดว้่า ในสภาวะที่
พืชได้รับความเข้มของแสงน้อย แสงจะเป็ นปัจจัยจ ากัดของกระบวนการสังเคราะห์ด้วยแสง แต่เมื่อความเข้มแสง
เพิ่มมากข้ึน อตัราการสังเคราะห์ดว้ยแสงของพืชกเ็พิ่มมากข้ึนดว้ยดงัที่ไดก้ล่าวมาแลว้ ดงัน้นั ในสภาพความเขม้
แสงมาก แสงไม่ใช่ปัจจัยจ ากัดอีกต่อไป แต่ความเข้มข้นของ CO2
จะเป็ นปัจจัยจ ากัดของพืช C3 ส่วนในพืช C4
พบว่าปัจจัยจ ากัดที่ส าคัญคือปริมาณเอนไซม์ที่เกี่ยวข้องกับการสังเคราะห์ด้วยแสง
ไม่มีความคิดเห็น:
แสดงความคิดเห็น